Hi, this year's Plant Genomes in a Changing Environment was virtual with hashtag #PlantGenomes20. Check the full program here. I post here my notes on the talks I was able to attend, either live or on the recorded videos, in case they're useful for anyone out there.
Giles Oldroyd (Sainbury lab, UK) talked about a pathway conserved in plants forming intracellular endosymbiotic mycorrhizaephytes. The pathway is not found in species forming exclusively extracellular symbioses, such as ectomycorrhizae, and those forming associations with cyanobacteria. This is described at https://www.nature.com/articles/s41477-020-0613-7. Receptors used with potential symbionts are the same used to recognize chitin and peptide-glicans of pathogens. Symbiotic potential decreases when there's enough nutriends at hand.

Figure from https://www.biorxiv.org/content/10.1101/804591v1.full
N Marmiroli (SITEIA.PARMA, Italy) presented a poster about the https://simbaproject.eu, where they are testing in plots in Italy and Germany the application of Plant Growth-Promoting Microbes to wheat, maize, tomato and potato under abiotic stress conditions.
Trevor Nolan (Duke U, USA) talked about their efforts in harmonizing single cell mRNA sequencing data from 110K cells to construct an atlas for the developing Arabidopsis thaliana root. Read more at https://www.biorxiv.org/content/10.1101/2020.06.29.178863v1
Francesco Licausi (Oxford U, UK) described the strategies of flooding tolerance in plants, which can be used to breed flooding varieties of crops. Flooding is a composite stress, he focus on two components: oxygen sensing and responses to oxidative stress, which are mediated among others by NAC transcription factors such as ANAC017, read more at https://onlinelibrary.wiley.com/doi/full/10.1111/pce.13037 and https://onlinelibrary.wiley.com/doi/abs/10.1111/tpj.14975
Ana Caño Delgado (CRAG, Spain) delivered a lecture on physiology and molecular biology of
Arabidopsis plants and their responses to drought mediated by brassinosteroids receptors in the root tip. Among many experiments, they measure response to drought as function of root branching angles. Read more at
https://www.nature.com/articles/s41467-018-06861-3 and
https://science.sciencemag.org/content/368/6488/266.abstract
Steve Long (Lancaster U, UK) shared his knowledge on breeding for incresing photosynthesis efficienty. This figure is from a 2010 review https://www.annualreviews.org/doi/10.1146/annurev-arplant-042809-112206, but still quite impressive:
Alexander Borowsky (U California, Riverside, USA) uses their own coexpression network pipeline (Wisecaver 2019) and the Taiji pipeline to analyze their ATACseq data in order to find key transcription factors regulating responses to water logging and water deficit in rice. They plan to do validation work using CRISPR to confirm their results.
Julia Bailey-Serres (U California, Riverside, USA) gave a keynote with two parts. While the second was related to A Borowsky0s talk, on the first part, she and her team looked at the transcriptional regulation of responses to submergence/flooding by comparing 4 different species (rice, Medicago truncatula, domesticated tomato and its dryland-adapted wild relative Solanum pennellii). By combining RNA-seq and ATACseq data and different computational analyses they found 68 Submerged Upregulated oRthologous Families (SURFs) upregulated in both monocots and dicots, with hypoxia-responsive promoter element (HRPE) motifs being abundant in rice but scarce in promoters of SURFs in dryland-adapted S. pennellii. This piece of work is described at https://science.sciencemag.org/content/365/6459/1291
Figure from https://science.sciencemag.org/content/365/6459/1291
They have really nive figure, shown below, where DNA motifs instead of transcription factors are the vertices of the graph. Despite the differences observed in the network across species, 4 motifs seem to be highly conserved:
Figure from https://science.sciencemag.org/content/365/6459/1291
Klaas Vandepoele (VIB-Ugent Center for Plant Systems Biology, Belgium) and his team have been doing a conceptually similar analysis, with RNA-seq from A. thaliana, Populus trichocarpa and maize, in order to identify gene triplets with conserved leaf expression pattern which are conserved across monocots and dicots. They validate their results by phenotyping A. thaliana mutants in the glass house. Their work has been published at https://onlinelibrary.wiley.com/doi/full/10.1111/pbi.13223. He also mentions this paper https://pubmed.ncbi.nlm.nih.gov/31748815 which describes modules of genes involved in cell proliferation in the leaf.
Jeff Habben (Corteva, USA) describes their extensive field trials in low and high-yielding locations, mostly across the US, to check whether different transgenic lines that overexpress maize transcription factor zmm28 (MADS-box) have higher and more stable yields. Their work is described in detail at https://www.pnas.org/content/116/47/23850
Facundo Romani (CONICET, Argentina) explains how liverwort Marchantia polymorpha defends itself against hervibores by accumulating terpenes in oil bodies, a process controlled by HD-ZIP transcription factor MpC1HDZ. This work is described at https://www.sciencedirect.com/science/article/abs/pii/S0960982220307685
Sandra Knapp (Natural History Museum-London, UK) , a plant taxonomist, talks about diversity of Solanaceae (nightshades), starting with potatoes.
Source: Sandra Knapp
She discusses how we usually think of aridification as something bad but according tho their work in Australia (https://onlinelibrary.wiley.com/doi/full/10.1111/jse.12638) it seems to be certainly a driver of diversification. She suggests we should be looking at plant lineages that have this pattern to prepare for climate change. One possible way would be to learn from resistant weeds in order to create resistant crops (https://www.sciencedirect.com/science/article/abs/pii/S0304423818307994). This is aligned with the work of Dana MacGregor, which also had a talk in this session.
She terminates by acknowledging the collective effort of people studying nightshades around the world (http://solanaceaesource.org) and a warning about the excess of P/N consum by agriculture and the current rates of extinction, that cause an unsustainable loss of diversity.
Michael Purugganan (New York University, USA) talks about the domestication of rice (which tokk 4K years) and molecular studies to understand the geographic spread of primarily japonica rice in Asia. His two main figures are:

Figure from https://pubmed.ncbi.nlm.nih.gov/28087768/
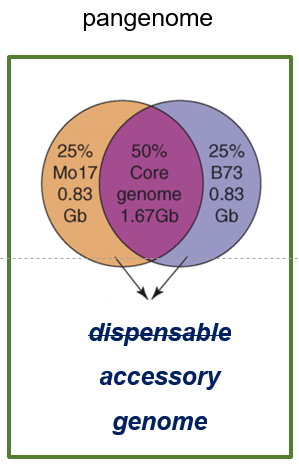


















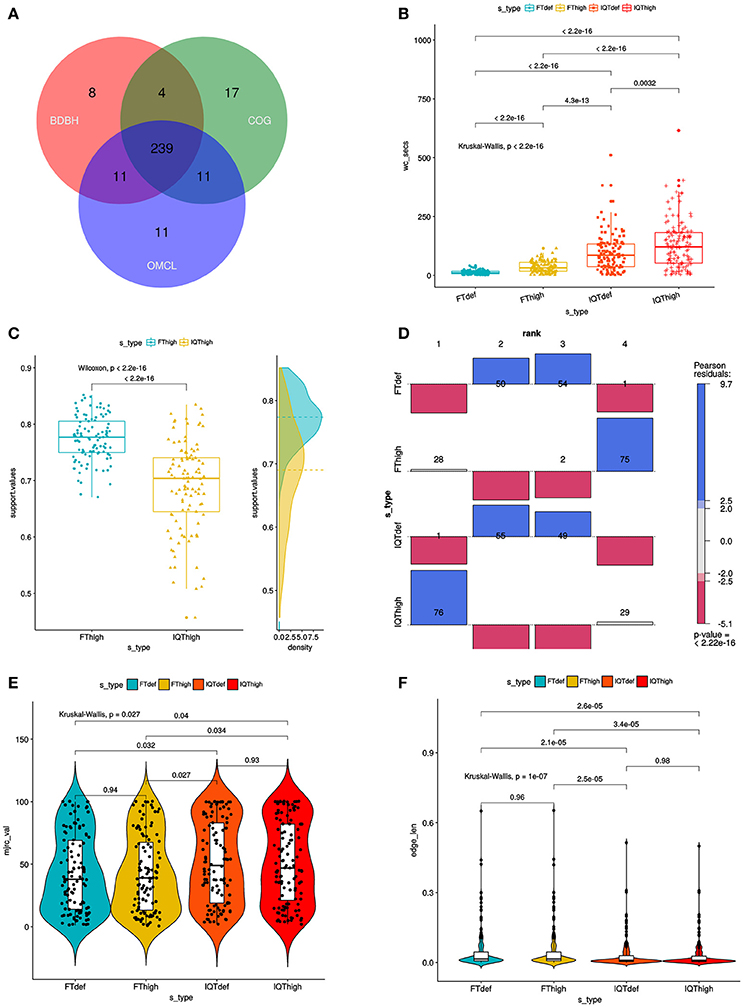{kind=link}